
 Recently, Freddie DeBoer published a book titled "Cult of the Smart" that outlines a hereditarian (somebody who believes in direct and isolated genetic causes determine complex behaviors) left-wing perspective on education. The central thesis of the book is that individuals come endowed with an "intrinsic ability" to excel in school. Some students, no matter how much effort is placed on improving their abilities, will fail, and it's therefore cruel to link ones economic prospects and social welfare to educational outcomes. As somebody on the left I am in broad agreement with decoupling academic from economic prospects, but when talking about the genetic influences over behavior Freddie writes in line with Plomin's recent flawed book "Blueprint" (see review here). One gets the feel that the author took a dive into popular accounts and blogs written by behavioral geneticists, but remained largely (willfully?) unaware that this whole field has been marred in controversy throughout its short existence. Large parts of the book opine that while we are told that our destiny can be the stars, in reality its deeply engrained in our genes. What's not discussed is the near failure of molecular methods to uncover this underlying genetic architecture for human behavior. As stated eloquently by Aaron Panofsky in his book "Misbehaving Science":
"Molecular genetics has been a major disappointment, if not an outright failure, in behavior genetics. Scientists have made many bold claims about genes for behavioral traits or mental disorders only to later retract them or to have them not replicated by other scientists. Further, the findings that have been confirmed, or not yet falsified, have been few, far be tween, and small in magnitude." As somebody who studies behavioral development it struck me how easily the author just accepted heritability estimates were an accurate measure of genetic causation without giving the scholarship on this topic its due diligence. This is surprising because the application of heritability estimates on humans has had a contentious history, and there is a large body of scholarly work on both sides to draw from. At its core heritability estimates are a population level descriptive statistic that calculates the recurrence of specific traits in families. They don’t measure genetic causes. Like any statistical measure it also comes with specific assumptions. In the case of heritability estimates, its core assumption is the existence of unmeasured direct and isolated genetic and environmental causes that exist at the individual level, and can be uncovered via partitioning phenotypic variation at the population level. While such partitioning has been a convenient way to bypass developmental processes, research on development has shown such partitioning to be a limited (and in most contexts, such as humans) invalid assumption (see https://doi.org/10.1080/10407410902877157 & https://doi.org/10.1111/j.1467-7687.2007.00556.x). This means that researchers who use heritability estimates often have much more nuanced perspective now than they did 10 years ago. Furthermore, the existence of "missing heritability" where heritability estimates from genomic techniques (such as GWAS and polygenic risk scores) are often smaller by orders of magnitude than heritability estimates from twin studies, has further questioned the ability to heritability studies to uncover genetic causes. While such molecular techniques are mentioned in the book their lack of concordance with more traditional behavioral genetics is glossed over completely. Freddie DeBoer also seems to take at face value a core assumption from behavior genetics the dose not hold up to scrutiny; the equal environments assumption (EEA). The EEA states that for all intensive purposes people sharing the same household have the same "environment". Many of the studies he cites have strong methodological problems. For example, in the Minnesota Twin Family Study monozygotic twins reared separately were often raised by relatives on the same street, interacted constantly, and went to the same school. These twins were however considered to have an "unshared environment" simply because they were raised in a different household. Controlled lab studies have also questioned the feasibility of the EEA. In 1999 a group of mouse geneticists across different labs tried to uncover the underlying genetic structure of behavior by standardizing the environment of inbred laboratory mouse strains across labs to a degree never achieved before. The result was large phenotypic differences both within and across labs, highlighting the stochasticity of developmental processes and largely putting the EEA to bed for everybody outside of a few behavioral geneticists (https://science.sciencemag.org/content/284/5420/1670). Further research has shown that even clonal mice housed in identical environments will develop distinct individualities based on slight social and stochastic differences in direct experiences (https://science.sciencemag.org/content/340/6133/756). The EEA does not hold. Freddie's book largely ignored the evidence from developmental psychobiologists, and throughout the book fails to address the near-consensus that the unmeasured developmental confounds outside the genome contribute to high heritability estimates in humans. He unquestioningly accepts the supposed fact that educational outcome are 0.5 to 0.8 heritable (which he interprets as upwards of 80% of educational outcome are determined via "genes") despite the fact that in controlled laboratory conditions, the only consistent heritability estimates for much simpler behaviors in animals are lower. The fact that even "purely genetic" conditions such as phenylketonuria (caused by a single nucleotide change in a single gene) can be cured with a 100% environment intervention (removing phenylalanine from the diet complex) should give one pause when discussing outcomes such as "education" and "intelligence". But not here. All behaviors investigated in detail have show that gene-organism-environment interactions form an irreducible network of ontogenetic resources necessary to cause the expression of any behavior at any point in time. There is, simply put, no room anymore for considering direct, isolated, & magnitude specific genetic or environmental causes within the interactive and relational reality of biological development. I ask Freddie DeBoer to read "The Ontogeny of Information" by Susan Oyama, "Synthesizing Nature-nurture" by Gilbert Gottlieb, "Basic Instinct" by Marc Blumberg , "The Dynamics of Behavioral Development" by Zing-Yang Kuo and "Design for a Life" by Patrick Bateson to balance out the one sided treatment drawn from the niche and controversy laden field of behavioral genetics. The book draws from a very limited range of behavioral genetic researchers, some of which who's work on the genetics of education have seen flattering endorsements from Charles Murray, author of the notorious and racist "Bell Curve". Many arguments in the book are supported by researchers, bloggers, and think tanks interested in "public choice" and market fundamentalist school policies that directly contrast with the progressive vision offered by the book. I urge Freddie DeBoer to take developmental research seriously in the next revision of the book (starting here: https://www.karger.com/Article/Abstract/73306) as it will both strengthen the case for progressive educational policy with real empirical findings about individuals rather than the stale, banal, and un-actionable application of heritability estimates to educational outcome.
0 Comments
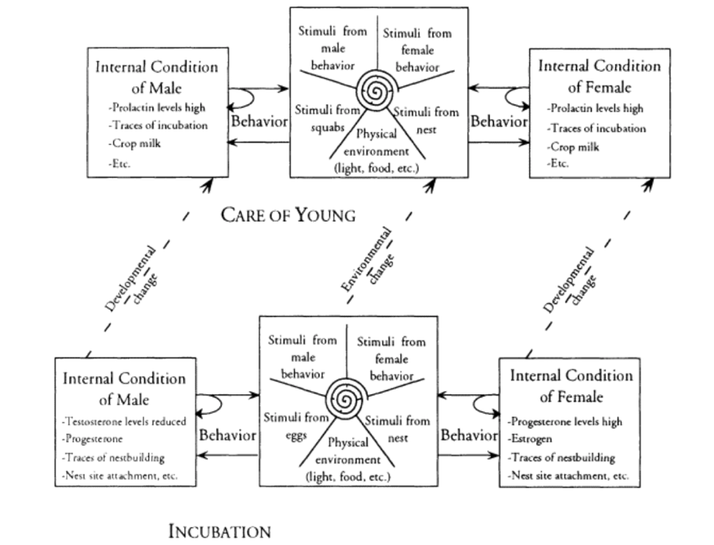 Fig 1. From Lehrman 1965 showing a network of contingencies shaping incubation and the care of young in ring doves. This highlights how the 'information' for these traits is distributed both within an organism and between an organism and its environment. In no individual part of this causal network is there the potential for a organizing developmental program, whereas the behavior emerge based on a recurrent sequence of antecedent-consequent interactions ultimately constructing ‘incubation' and ‘care of young’. Fig 1. From Lehrman 1965 showing a network of contingencies shaping incubation and the care of young in ring doves. This highlights how the 'information' for these traits is distributed both within an organism and between an organism and its environment. In no individual part of this causal network is there the potential for a organizing developmental program, whereas the behavior emerge based on a recurrent sequence of antecedent-consequent interactions ultimately constructing ‘incubation' and ‘care of young’. How biology constrains, shapes, and facilitates human potential has always been contested territory, and nowhere has this debate been more contested than in discussions of human 'nature'. While the term 'human nature' may seem like an antiquated throwback to the days of vital essences and lumeniferious aether, I have been continually surprised to see individuals citing the modern denial of our 'nature' as the root cause of many societal ills. What's the basis of such concerns, and why do so many people put so much importance on a seemingly ill defined abstraction such as human 'nature'? I believe the importance given to human nature in our public discourse in part reflects the way that researchers approach how evolutionary processes influence our everyday lives. At our “core” are we just objectified manifestations of selective forces that acted on generations of humanity before us, or is each individual a novel synthesis of past evolutionary factors and here-and-now factors. The science that has attempted to answer these questions has changed significantly over the years, from sociobiology, to human ethology, to human behavioral ecology and evolutionary psychology. According to evolutionary psychology our everyday lives are shaped to varying degrees by a series of evolved psychological and cognitive programs/modules. It is hypothesized that these programs subtly shape nearly every aspect of our lives, from our mate choices, to the products we buy, to the political ideologies we adopt. Furthermore, these programs have such a large influence on us because they represent adaptations that have been instantiated in our biology by generations of natural selection. Despite being in the business of studying adaptations, defenders of evolutionary psychology rightly go to great lengths to resist claims that they are pan-adaptationists by proposing that psychological processes can be organized into three bins: adaptations, byproducts, and noise. A main assumptions is that one can differentiate psychological adaptations from byproducts or noise by documenting that specific traits have prespecified purpose and universality. This assumption provides the foundation for most hypotheses in modern evolutionary psychology. I argue that not only are such strong demarcations between adaptations, byproducts, and noise unnecessary for evolutionary processes, but that common approaches to estimate such demarcations are unavailable to evolutionary psychologists, and students of human behavior more broadly. Linking adaptations with shared functionality and universality has it’s roots in early ethology. One of the major advances that ethology introduced to the study of behavior was the ability to view behavior from an evolutionary lens. Much like a taxonomist could reconstruct the evolutionary relationships between species by comparing their morphologies, ethologists showed that one could do the same by comparing the behavior across different species. This approach was exemplified by Konrad Lorenz in his excellent book "Comparative Studies on the Behaviour of Anatinae” that built upon his advisors Oskar Heinroth’s studies of the evolution of behavioral displays in ducks and geese. Nonetheless, the early ethologists also appreciated the variability of animal behavior, and tended to only apply evolutionary analysis only to behaviors that met a certain criterion, they had to be universal, or develop predictably across all members of a species or population, and they had to have a purpose, they allowed the organisms to correctly respond to environmental challenges they had not yet experienced. These behaviors, called modal or fixed action patterns, were treated as the products of discrete developmental programs that assured that specific behaviors developed independent of specific experiences. Thus, as a healthy member of species X you would display functional behavior Y in response to some stimuli independent of your specific experiences with your environment. Until Lehrman's critique the main way that ethologists identified modal action patterns was to raise an individual in deprived settings to show that some behavioral traits and abilities were still predictably expressed. Once modal action patterns were identified comparisons between species could be made to investigate their evolutionary history. Much of the discussion around evolutionary psychology rests on its treatment of specific psychological and cognitive tendencies as equivalent to modal action patterns. Adaptive psychological and cognitive processes must be universal and purposeful, as stated by Dr. Al-Shawaf this means that "psychology and behavior predict universality at the level of the information-processing structure of the neurocognitive mechanisms that produce behavior”. This universality suggests that these psychological abilities arise independent of an individual’s specific experiences, and represent an environment than an individual has not yet, or may never, encounter (often proposed as the environment of evolutionary adaptedness). This requires that development is teleological, that it's directed towards specific outcomes through the utilization of developmental programs that unfold specific functional traits at the right time during ontogney. In response to similar claims made by ethologists many American comparative psychologists, such as T.C. Schneirla, Daniel Lehrman, Frank Beach and especially Zing-Yang Kuo, questioned the independence and preparedness of modal action patterns. In particular, research done by these comparative psychologists showed how behavior that seemed prepared for future environments were often developmental variations of prior functional behaviors utilized in novel ontogenetic contexts. For example, walking and gait in hatchling chickens were developmental reorganizations of similar leg movements made pre-natally to stimulate respiration and proper positioning in the egg. In particular they showed that even in isolation studies, specific experiences between an organism and its environment were essential factors in shaping the emergence of modal action patterns over development. Since then experiments have consistently demonstrated that the development of modal action patterns is dependent on a network of interacting developmental resources extending both outside and inside organisms. Therefore, the information “for” modal action patterns was not latent within the organism or its genes, but constructed via contingent relationships within and between an organism and its environment (Fig 1.). As stated by Oyama (1988) "the informational function of any developmental interactant is dependent on the rest of the system. This means that what counts as information is itself contingent and relational, not that information “is” the developmental system.” Thus, if latent developmental programs exist for specific behaviors it is unclear how they could be instantiated without postulating some near supernatural force sitting outside both the organism and its environment and causally guiding and coordinating all the relationships within and between them. In response to these findings’ ethologists such as S.A. Barnett, Patrick Bateson and Johan Bolhuis began to question the assumption that modal action patterns occurred independent of specific experiences and proceeded to create a developmentally informed ethology without “instincts”. A uniting theme behind this new ethology/comparative psychology was that development was re-conceptualized as emerging prospectively. That is, all behavior arises from earlier conditions, but shows no direction towards later conditions. Development was freed from its teleological constraints. The behavior of any organism at any point in time reflected the current state of the organism and its relationship with its environment, and these antecedent states provided the foundation from which other consequential states can arise. There was no need for any program, guide, blueprint, invisible hand, vital essence or other abstraction to guide development to a final endpoint, as the lifecycle of organisms was simply a series of antecedent-consequent processes that reoccur anew within each individual. In contrast to this prospective view many behavioral researchers took a retrospective view that grounded developmental questions upon specific adaptive traits of interest. Here one looks back in time, starting with the trait under study and looks for potential developmental causes arising earlier in life. Such a teleological view often masks important developmental processes by selecting only those traits early in development that share some functional or perceptual similarity with the trait of interest (In contrast, the prospective developmental approach has continually shown the relevance of non-obvious and non-linear experiences in the development of modal action patterns). The fixivity of specific behaviors over ontogeny, or the seeming directionality of their development—which are a consequence of the retrospective perspective and not development per say—is often seen as evidence a developmental program, and used as a stand-in for past selection pressures. According to this view the development of adaptive traits don’t so much emerge as a consequence of previous states but unfolds according to an evolved program. Such developmental programs have often been used in evolutionary psychology as a way to assure that adaptive traits maintain their universality. This assumption has been sustained despite the lack of evidence that biological or psychological processes function like a program. In recent decades there has been a steady erosion of colloquial assumptions that genes function as developmental blueprints or programs, or that brains only act as computational programs that take in stimuli as input, process sensory information, and spit out behavior as outputs (This has its roots in Dewey’s classic 1896 critique of the reflex arc). This leaves one with the difficult question of where developmental programs are instantiated in the inherently interactive network of factors that define all psychological events. If one is unable to provide a testable and measurable substrate from which a program could be instantiated, it might be better to assume the more empirically supported observation that the universality of specific traits represents the shared antecedent-consequent interactions reconstructed across individual lifecycles. So what does this have to do with evolutionary psychology? Since phylogeny is just a reflection of recurrent and successful lifecycles, and those lifecycles consist of recurrent antecedent-consequent processes, there are opportunities for sudden, universal, and functional novel phenotypes to emerge via developmental reorganization. As stated by Lickliter 2014, "The novelty-generating aspects of development involved in such evolutionary change are the result of the developmental dynamics of living organisms, situated and competing in specific ecological contexts, and not simply the result of random genetic mutations". This process, called neophenogenesis, lays at the heart of the difficulties in demarcating where adaptive phenotypes begin and end. Many cases of neophenogenesis are considered noise, single individuals whom were shaped by incidental exposure to some experiences over development. Such events have now been widely observed, but “ noisy” responses can also cause animals to flexibly reorganize their developmental resources to solve the here-and-now challenges of everyday life. This has been seen in many studies of animal innovation. In many cases of animal innovation the behavior emerges and dies within a single individual, but it is increasingly shown that such innovations can spread to higher levels such as the group, population, and potentially the species. Thus identifying a shared functional behavior does not automatically mean that the behavior is the cumulative result of generations of selective pressure by default, as an increasing number of studies (especially in birds and mammals) have shown that novel, adaptive phenotypes can arise and spread quickly across a population. Large scale environmental changes can induce coordinated changes in developmental processes. Group and population level developmental accommodations have been seen in invasive species, ontogenetic responses anthropogenic changes, and responses to changes in social dynamics. Such examples highlight the ability reorganize developmental processes to coordinate changes across a whole group or population by exposing all individuals to novel shared experiences. Often these shared developmental accommodations can be directly functional, allowing individuals to survive and reproduce in the new novel environment. This has been observed in population scale shifts in migratory routes, mating patterns, and egg clutch size in European blackcaps exposed to sudden anthropogenic changes (Bearhop, S, et. al. 2005) and population level phenotypic divergence mediated by maternal effects in House finches moving into novel habitats (Badyaev, 2005). Another example of this can be seen in Terkel's studies of pine cone foraging in Israeli black rats. Throughout most of their range black rats do not consume pine nuts. However, a whole population of black rats learned to harvest pine nuts through developmental accommodations in a plantation of Jerusalem Pine in Israel (Fig 2). Here a single individual likely learned to open a pine cones themselves, and this spread throughout the population through the introduction of a new developmental resource, partially opened pine cones. Juveniles exposed to the partially opened pinecones left by adults were able to acquire the ability to open cones on their own over ontogeny thus constructing a shared functional developmental accommodation. Such developmental accommodations make it difficult with limited observations on a single population or species to assess if behavior reflects the refined product of generations of natural selection, or shared developmental accommodations meant to address here-and-now challenges. The whole dichotomy between an adaptation and a developmental accommodation becomes blurred, as all aspects of an organisms represent a synthesis between the past and present. This synthesis is necessary as all organisms must navigate an environment (either ecological or perceptual) that is a shifting mosaic of regularities and novelties. Even if one conceptualizes an evolved psychology as nothing more than a series of adaptive modules, the connections between the utilization of different modules and novel environment challenges would continually construct novel information processing networks that may bear little resemblance to their ancestral function.  Fig 2. Likely a single innovation, a rat learning to open a pine cone quickly spread throughout a whole population of Israeli black rats. This novel developmental accommodation was sustained across the population as young individuals predictably encountered the half-opened pine cones from adults, thus recreating the acquired behavioral pattern anew during each lifecycle. Here lies the issue for evolutionary psychology. Their hypotheses relies on a demarcation between adaptation and byproduct through identifying universality and functionality of specific psychological abilities. A common response among evolutionary psychologists when faced with the emergence of functional neophenotypes is to claim these are simply the flexible outputs of the adaptive underlying cognitive-informational processing programs. However, this simply kicks the problem down the road by instantiating the program at a lower level of biological organization. Nonetheless, no level of biological organization is immune to developmental contingencies or accommodations. Studies across all levels of biological organization, from gene expression to social organization, have documented the routine occurrence of shared and functional developmental accommodations. It is therefore unclear that even if one could locate where a universal adaptive program is instantiated, that it would be immune to the effects of developmental accommodations. Another issue for evolutionary psychologists are developmental accommodations itself. Humans unlike most animals don’t have many (or any, depending on whom you ask) modal action patterns. However, the production of shared and functional developmental accommodations is what humans do, all the time, continually, on a scale and magnitude not seen before in any other species. Large scale changes in organismal-environment relationships that continually create novel brains, bodies, biologies and behaviors is part in parcel to human history. Human beings have are masters at manipulating the antecedent-consequent structure of developmental processes to directly create adaptive neophenotypes outside of, and in front of, selective forces. At no point in human history, be it the Pleistocene, the bronze age, the middle ages ,or the information age, can one stop, put a stake in the ground, and demarcate a line between conserved selective features and novel developmental accommodations. Humans are ultimate developmental niche constructors, we have the ability to plan, understand, and expand our potential by radically reorganizing our biology and behavior. Our potentials vasty outweighs the constraints of our ancestors. So what's the way forward? One way is to make evolutionary psychology truly evolutionary. A recent blog post by Kevin Bird discusses how evolutionary psychologists should apply techniques from evolutionary biology to identify where selection has, and is, acting. Nonetheless, without a detailed historical record or closely related comparison species it is woefully difficult with the current methodologies used in modern evolutionary psychology to identify phylogenetic history and selection. Another approach is to fully integrate the production of developmental accommodations into the study of evolutionary psychology. Baldwin (1896), Osborne (1986) and Loyd Morgan (1896) proposed testable hypotheses regarding how developmental accommodations and neophenotypes facilitate natural selection. Broadly, the continued production of functional neophenotypes is first maintained cross generations via recurrent developmental accommodations. Selection acts on these recurrent accommodations by selecting ontogenetic processes that more efficiently and predictably produce the trait under selection. This process may lower the energetic burden of producing complex traits over development, thus "freeing" the organisms to develop new accommodations in response to changing environments. This "assimilate-and-stretch" model outlined in Avital and Jablonka (2000) suggests that the continual production and assimilation of neophenotypes is a core component in the evolution of behavioral complexity, and continually blurs the line between adaptation and neophenotype. Niche construction theory proposes that organisms possess the ability to shape their own selective pressures through their influences on the external environment. Rather than just assimilating developmental accommodations in response to external pressures, the feedback between the emergence of neophenotypes and their corresponding changes in the environments creates novel selective landscapes. From the evolution of lactase persistence, to the emergence of agriculture, to the development of livestock domestication, and the continuing use of technology to drastically alter our psychologies and biologies there is little reason to doubt that niche construction has shaped the evolutionary trajectory of our species. Instead of attempting to identify signatures of past adaptions, evolutionary psychologists could be investigating at how novel interactions and feedback loops between human behavior and the environment could shape the continuing evolution of our psychological abilities. In conclusion, the fact that developmental accommodations can create widespread, novel, and functional phenotypes in front of selective pressures provides a large obstacle for our ability to understand the selective forces shaping human behavior. However, approaches emphasizing the role of developmental systems in evolution (as outlined by Johnston and Gottlieb (1990)) may inform a more robust and evolutionary approach to researching human behavior by focusing on which traits will lead us forward, rather than how the past constrains us. 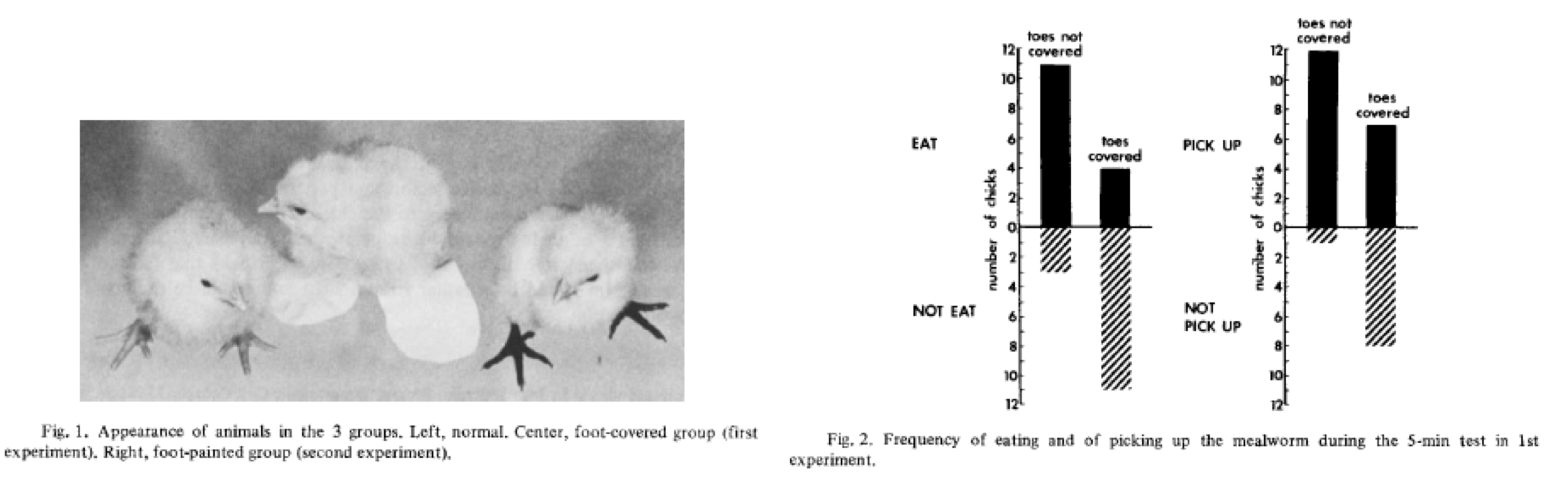 “Nature is probably quite indifferent to the aesthetic preferences of mathematicians.” ― Alfred North Whitehead The tendency to assume that we can understand how complex psychological processes work with a simple equation, or even a single number, is seductive. Heritability is an attempt to describe the processes wherein traits are inherited across generations with a single number. The use of heritability is appealing, because it reflects many peoples common sense knowledge about inheritance —that behaviors, abilities, and traits tend to be more similar within families than between them. However, common sense is an awful guide for scientific investigation, especially developmental science. In developing organisms many non-intuitive factors play critical causal roles in the development of specific traits. One such example is that baby chicks require the experience of seeing their toes to learn to peck and consume mealworms (Wallman, 1979). In such cases, it is highly unlikely that a prior and targeted hypotheses would be able to pinpoint all the necessary and sufficient factors for the development of that trait. Studying development is hard, it involves lots of longitudinal observations, persistence, grit, trial-and-error, and a fair amount of luck. Those willing to spend the time observing organisms across development can be handsomely rewarded, often by discovering never-before-seen factors in the development of behavior that can then be investigated using targeted experimental approaches. Faced with the difficulties of researching development, it is tempting to go a simpler route, by ignoring the details of individual development and assuming that traits are exchanged between generations in “units” of information—genes that bias organisms towards aquiring some traits and not others. Behavioral and population geneticists often never directly measure genetic contributions, but estimate them by measuring how variation in traits across individuals is predicted by sharing genes with others (comparing related individuals or monozygotic twins) or sharing environments with others. By separating the variance in terms of genetic and environmental components they can calculate a single number that show how much genes control the development of a trait. A heritability estimate of 0.0 means that a trait is entirely controlled by the environment and a heritability estimate of 1.0 means that a trait is entirely controlled by genes. One of the main disadvantages of heritability estimates is that they attempt to calculate the genetic contributions underlying traits in a population by reducing the complex process of individual development. It does this by assuming that all traits have an independent genetic and environmental component (and sometimes and interactions between the two) during development. In animal breeding operations where husbandry practices carefully control many aspects of the developmental environment heritability estimates have some utility, although even in these conditions they do not tell you anything about the actual processes underlying the ontogeny of traits of interest. For example, one could easily the calculate heritability estimates on early responses to mealworms in chicks, but it would never tell you that chicks need to see their own feet to develop the ability to peck and eat mealworms. Developmental denialism occurs when heritability is treated as a causal mechanism governing the developmental reoccurrence of traits across generations in individuals. Such an approach thwarts true developmental science by assuming a priori which mechanisms are causal in the development of specific traits. Opposition to heritability as a causal explanation is not new. Zing-Yang Kuo (1929) was arguing for the abolishment of heritability in psychology in the 1920s, and afterwards, American comparative psychologists T.C. Schneirla, Gilbert Gottlieb, and Ethel Tobach argued for a developmental approach to the reoccurrence of traits. Why is developmental denialism a problem? Because researching development is the only way to really understand where differences and similarities between individuals come from, it is the best way to identify causal factors shaping the emergence of differences, and design interventions to correct differences that lead to disadvantages. Assuming that highly heritable traits are “mostly genetic” thwarts research into understanding the actual mechanisms governing the emergence of traits, it creates black boxes in lieu of understanding how genes and environments shape development, and provides just enough empirical clout to be misused by those with nefarious political aims. In particular, three major issues with using heritability to deny development should be further elaborated. Issues with using heritability as a replacement for development: 1. Genetic and environmental influences are never independent 2. Analysis of variance can not uncover causal developmental processes 3. The organism is negated It often surprises people when I say that I believe everything is genetic, because genes are a continually present factor shaping organisms throughout their development, and without them there could be no development at all. But the same could be said for a wide range of different developmental resources. Try developing without mitochondria, gravity, light, water, food, or interactions with other individuals. All these factors are essential for an organism to maintain and perpetuate itself. Furthermore, variation in these factors will shape variation in phenotypes. But genes do not occupy a privileged, centralized role in development, they do not direct, control, or guide development any more than other factors. This is because the information in genes is always relational, they are never independent of other factors, and do not have a direct, isolated, or linear influence on organisms. For example, the gene for FoxP2 protein plays a role in vocal learning in the brain, liver function, and even cancer based on differences in where, and how, the gene is used by the organism during development. The information in genes is dependent on the state of the organism, its behavior, its environment, and its developmental stage. Thus a core assumption of hereditarianism, that organisms can be partitioned off into separate independent environmental and genetic influences denies the reality of how development actually works. Heritability, and analysis of variance more broadly, can not identify causal developmental processes. Moore (2006) discussed this in his paper on the heritability of IQ, and I will repeat his argument here. Snow only occurs when temperature and moisture are at the correct levels. If one was to use an analysis of variance to look at the causal factors underlying the presence of snow in montane tropical environments and Antarctica you would get very different results. In Antarctica, where the temperature is always low enough for snow to occur, only variation in humidity that would correspond with the presence of snow, thus humidity would seem to “cause" snow. In montane tropical forests where humidity is always high it would be differences in temperature that lead to the presence of snow, thus temperature would seem to “cause" snow. Using heritability estimates to uncover independent causal processes is misguided, as multiple factors can shape variation in traits across individuals in the population, and the heritability estimates will never be able to uncover the processes that generate such variation across individuals. Another problem of this approach is that traits that don't vary across individuals are always environmental. For instance, the ability to use language is a uniquely human ability. Since linguistic ability is ubiquitous across all human populations, and thus does not vary, by calculating a heritability estimate of linguistic ability you will come to the conclusion that it is entirely environmental. This would directly contradict the established literature as the development of language has been shown to be the emergent consequence of growing up in a linguistic social environment coupled with unique neurogenic mechanism that allows children to quickly develop language during early life. My largest issue with hereditarianism is that it negates its own object of study, the organism itself. By treating organisms as bundles of independent traits each separated into independent genetic and environmental influences, it ignores the fact that the organism is a whole integrated system that continually interacts and changes its proximal environment throughout its life. The organism is a nexus where genes and environment interact, it is an irreducible common reference point for all of biology and psychology. The behavior of the organism can feedback into shaping both the environment it experiences and the expression and functionality of the genome across ontogeny. When taking the development of the organism seriously it quickly becomes apparent that causality is bidirectional going from genes-to-organism-to-environment as easily as it can go from environments-to-organism-to-genes. Wallman, J. (1979). A minimal visual restriction experiment: Preventing chicks from seeing their feet affects later responses to mealworms. Developmental Psychobiology: The Journal of the International Society for Developmental Psychobiology, 12(4), 391-397. Moore, D. S. (2006). A very little bit of knowledge: Re-evaluating the meaning of the heritability of IQ. Human Development, 49, 347 – 353. Kuo, Z. Y. (1929). The net result of the anti-heredity movement in psychology. Psychological Review, 36(3), 181. Theoretical biologist Paul Weiss started his introductory biology courses by presenting two test tubes to the class. One test tube contained viable embryos and the other contained embryos after homogenization (basically blending them). He asked the class why, despite having the exact same chemical constituents, the viable embryos would be considered living, while the other test tube wasn’t. This exercise was meant to highlight the absurdity of reducing organisms to just their chemical constituents, and that interactive and combinatorial processes were more important to understanding organisms than simply knowing what they were “made of”. Today, esteemed biologists such as Jerry Coyne seem to be moving in the opposite direction, suggesting that organisms and behavior can be understood as outcomes of the chemical compositions of our cells and genes. Bioinformatics approaches to biology such as Genome-Wide Association Studies and RNAseq seek to link phenotypic traits with the presence or amount of molecular “components” in specific tissues.
Such a strong reductionist perspective cannot provide a complete picture of organisms. Most of us have spent time as children playing with Lego blocks. Like many biological processes, the interactions between molecules scale up to build larger structures, just as Lego blocks can be combined to make houses, boats, animals or other structures. The shape of Lego blocks prevents the formation of specific structures, as it's impossible to make a perfect sphere with rectangular blocks. You could even compute all the potential structures that can be built with a series of blocks, by modeling all the ways those blocks could connect to each other. Nonetheless, even with this knowledge we still have a very limited understanding of what actually will get built in real life situations. A complete catalog of all the Legos in a toy basket won't allow you to predict what structure a child will build. In order to understand the final structure, you need to understand the processes where the blocks are placed together. Thus, having a catalog of blocks allows you to calculate all the potential structures that could be built, but it can't predict which structures actually are built (the set of components defines the boundaries but not the specific outcomes). Many biologists, neuroscientists, and psychologists don't just seek to catalog the material components that make up organisms but to uncover mechanisms that capture how those components interact to influence the functional properties of organisms. Unlike Legos, the molecules in living systems are reactive. Entire research careers are often centered on a series of specific genes or molecules and how they react with other molecules to form physiological pathways that have functional outcomes. For instance, the gene VLDLR plays a critical role in the reelin pathway that is involved in cell-to-cell cohesion, neural migration, and diseases such as atherosclerosis. The assumption of such research programs is that a full catalog of all the mechanisms associated with some functional property, such as hunger, will lead to a complete understanding of that property. Such an assumption is called micro-determinacy where the interaction between the parts scales up to functional organism-level outcomes. Here causality is a one-way street from molecules to the organism. While mechanistic studies have been essential to uncovering key molecular pathways involved in nearly all aspects of living systems—from hormonal regulation of behavior to the neurogeneitc influences on learning, to trans-generational epigenetic inheritance, and development of novel medicines – the assumption that a complete catalog of mechanisms will allow for a complete understanding of living systems is coming under increased criticism. Recently, there have been attempts to test the effectiveness of such mechanistic approaches by applying standard methodologies from the biological and neurological sciences to human-engineered objects, such as a microprocessor running a video game or a radio. These studies test how effective our methods are at uncovering a “complete” understanding of the system under investigation. For instance, a common technique in behavioral neuroscience and genetics is to “knockout” specific brain regions or genes and observe their influence on the phenotype. Similarly, you can do a “knockout” experiment on electronic devices by removing components and observing changes in how the device functions. The results from such studies are sobering. In none of the studies were the approaches ever able to reverse engineer the system. Such findings highlight some real limitations of reductionist attempts to understand complex systems. Such criticisms do not take away from the obvious and widespread advances that such reductionist endeavors have achieved, but rather highlight how new concepts and methodologies might help to integrate experimental findings across levels into a less fragmented, more holistic, understanding of living systems. So where do we go from here? I propose that we start viewing organisms not as a static object but as a lifecycle. A lifecycle is a developmental system that is initiated at conception and flows until death. Organisms have little in common with Lego structures, radios, or computers. A defining feature of organisms is that they are thermodynamically unstable open systems that build and maintain themselves over time through a continual exchange of materials with the environment (also known as autopoiesis). From the moment of birth (or hatching, ect..) organisms are fighting for their lives. As highly organized systems, organisms must constantly engage with their environments to maintain their integrity. They must eat, drink, and breathe, in order to get the raw environmental materials to build and maintain themselves. Such materials are almost never uniformly distributed in the environment and usually occur in patches or gradients. This means that organisms must be active, by moving and interacting with their environment in order to secure and utilize the resources needed to persist over time. A plant grown in a dark room must take advantage of light gradients and grow toward high light levels to survive. The newborn rat must recognize and move toward their mother's nipple to secure milk. Even seemingly sessile organisms, such as barnacles must be active, by extending their filter like cirri into open water during periods when food is available. In general, there are two ways that all organisms can interact with their environments: they can either approach specific environmental stimulation, or they can withdraw from it. The comparative psychologist T. C. Schneirla was the first scientist to recognize this “biphasic process ofapproach-withdrawal” in his now classic 1959 paper, “An evolutionary and developmental theory of biphasic processes underlying approach and withdrawal.” Here, he outlined the basis of his comparative psychology, that developmental processes largely result from a refinement of approach/withdrawal behaviors that seek to perpetuate an organism's lifecycle, allowing them to grow, sustain themselves and reproduce (create new lifecycles). Organisms should approach and utilize resources in the environment that progresses their lifecycle, and avoid aspects of the environment that will work against the continuation of their own, or their offsprings lifecycle. [[ to be continued in part 2 ]] |
